その32 2013.3.9
このところずっとオオバキスミレのルーツ(Chamaemelanium節)を訪ねてインターネットの世界を探訪して来ました。先々月には
「探訪記31」であまり情報のない地下茎や閉鎖花に注目して近隣のアジアや北米の親戚達を紹介しました。その過程で興味深い論文を見つけたのです。ス
ウェーデンのヨーテボリ大学のマルクーセン(Thomas Marcussen)さん達が2011年に発表した「Establishing the
phylogenetic origin, history, and age of the narrow
endemic Viola guadalupensis (Violaceae)」という論文です。「狭固有Viola guadalupensis
(スミレ科)の系統学的起源、歴史および年代の確立」というほどの意味になるでしょうか。
http://www.amjbot.org/content/98/12/1978.longこ
こで全文を読むことが出来ます。
Viola
guadalupensisというスミレは1990年にテキサス州西部のグアダルーペ山脈国立公園の高山帯の石灰岩の岩場で発見された新種のスミレで、3
年前の「探訪
記30」でChamaemelanium節のリストを作成したときには私はまだその存在を知らなかったオオバキスミレの親戚の一員でした。
http://www.nwf.org/News-and-Magazines/National-Wildlife/News-and-Views/Archives/2010/Hidden-In-Plain-Sight.aspxこ
れは2010年の再発見を報じた雑誌記事ですが、魅力的な黄色い花の写真も載っています。
この論文はViola
guadalupensisに焦点を当ててはいますが、この種が属する北アメリカ全体のChamaemelanium節のメンバーの類縁関係や由来を考察
することもテーマになっていて、アジアのV. unifloraやわがV.
brevistipulata(オオバキスミレ)も材料に加えられています。オオバキスミレも含めたChamaemelanium節の分子系統学による研
究論文とあればもう読むしかありません。苦手な英語の、しかも専門用語が目白押しではあってもここで引き下がっては男が廃ります。幸いにしてインターネッ
トには外国語の文章の簡易翻訳機能がありますし、モニター上で単語を一つ一つ取り出して翻訳することも出来ます。悪戦苦闘の末ある程度の日本文にしまし
た。
最初の抜粋の部分は以下のように訳してみました。
•
研究の前提:気候の変化および土地利用の変更は生物多様性に対する2つの主な脅威である。そして狭固有種にとってはなおさら不均衡なインパクトになるであ
ろう。土地の管理者は、それらの起源および遺伝的多様性の程度を理解することによって、これらのユニークで高度に局所的な遺伝子プールをよりよく保存する
ことができるはずである。Viola guadalupensisはGuadalupe Mountain(北米テキ
サス州西部)の狭固有種である。Viola属Chamaemelanium節内の類縁関係はいくつかの議論の主題となっていた。更に、この節内の倍数体お
よび推測される網状の関係は大部分は未知のままである。
• 方法:我々は、V.
guadalupensisの染色体を数えた。また、24のViola分類群によって葉緑体trnL-F領域および低コピーの核遺伝子GPIの系統樹を生
成し、倍数体の系統ネットワーク構築に使用された。分岐年代は化石による較正によって得られた。
• 重要な結果:減数分裂における染色体数は、V. guadalupensisが4倍体(n=12)であることを明らかにした。また、2つのGPI
同祖遺伝子群の存在はさらにそれが異質4倍体であることを示唆した。系統関係の復元は、Canadenses亜節(父親)およびNuttallianae
亜節(母親)の未確認のメンバー間の交雑によってそれが起源したことを示した。
化石で較正されたGPI分析によるおおよその時間は8.6(5.7~11.6)Myr(=100万年)であると見積もった。これはV.
guadalupensisの最大 の年代が、Guadalupe
Mountainが1200~1300万年前に形成した後に進化したことを示唆している。
• 結論:Viola
guadalupensisは倍数化を伴う亜節間の交雑によって起源した。Chamaemelanium節の中では、この現象が、最後の900万年に(少
なくともV. bakeri、V. douglasii、V. glabellaおよびV.
sempervirensでは)繰り返し生じた。節の系統学的帰結が議論される。
以上、翻訳終わり。
マルクーセンさん達はViola
guadalupensisなどスミレ属25種を用いてDNAによる系統樹を作りました。その内訳はChamaemelanium節の7つの亜節の内6つ
をカバーする16種。この中にV. uniflora(ロシア産)とV. brevistipulata(日本産の栽培品オオバキスミレ)、そしてV.
biflora(ノルウェイ産キバナノコマノツメ)も含まれます。参照種としてその他の節から5種、そして作られた系統樹に根をつけるための外群として南
米チリ産のスミレ属4種です。研究に用いたDNAは核のGPI(グルコース-6-リン酸イソメラーゼ)をコードする領域と、葉緑体の遺伝子間領域
(trnL-F)の二つです。
DNAの塩基配列の変異を比べることで植物の系統関係を知ることが出来ます。母系遺伝する葉緑体DNAのなかでもtrnL-F領域は変異が大きいので近縁
種や種内分類群の系統解析に広く用いられてきました。一方核のGPI領域は両親から遺伝するので雑種由来の異質倍数体であればそれぞれの祖先から受け継い
だ遺伝子を複数持っていて、それらを系統解析することで由来を明らかに出来る可能性があります。
マルクーセンさん達は、両方の解析からChamaemelanium節の2倍体10種については、今までの形態的特徴に基づいて建てられた6亜節の分類
通りに纏まるけれども、V. guadalupensisなど6種の倍数体は、V.
canadensisが同じ亜節の種同士の雑種由来と想定される以外は、いずれも異なる亜節の種同士の雑種由来と推定され、いままでの亜節分類は成り立た
ないとしています。またChamaemelanium節のもっと多くの種を解析すれば、V.
guadalupensisなどの両親が分かるかもしれないが、すでに絶滅した種かもしれないとも述べています。
今回の探訪記では、前回の探訪記でも注目したV. uniflora(2倍体)とV.
brevistipulata(2倍体)、及び北米のV.
glabella(4倍体)とV.
pubescens(2倍体)、それとこの論文においてもChamaemelanium節に含まれるBiflorae亜節の一員として分析に加えられてい
るV. biflora(2倍体)も合わせて注目してみます。
まず、trnL-F領域による系統樹ではChamaemelanium節の16種は大きく二つのグループに分かれ、V.
pubescensなど北米の13種は一つのクレード(共通の祖先を持つ単系統群)に纏まります。しかしV.
brevistipulataなどのユーラシアの3種はChamaemelanium節以外の参照種の一種と合わさって一つのクレードとなっていて、
Chamaemelanium節だけでは単系統とはなっていません。そして3種の中ではV. unifloraとV.
bifloraがより近縁ということになっています。
一方GPI領域ではChamaemelanium節の遺伝子と参照種の遺伝子が入り乱れた系統樹をなしています。これは過去に南米から分布を拡げてきた
Chamaemelanium節の祖先が北半球で適応放散する過程で他のViola属のメンバーと幅広く交雑した歴史を反映しているらしいのです。しかし
北米の5つの亜節ごとに遺伝子群としては概ね纏まったクレードをなしています。ただV. rotundifoliaだけはどういう訳かV.
unifloraと単系統となり、V. bifloraはここではV. brevistipulataと単系統をなしています。
どちらの系統樹からもChamaemelanium節のなかで、V.
brevistipulataなどのユーラシアの3種はそれだけでは単系統群とはなりませんが、V.
pubescensなど北米の種とよりは近しい関係にあるのが見て取れます。
ここにきて、私も想定を改めなければならないと思いました。V. bifloraは柱頭や花弁の形、全体的な姿も、他のChamaemelanium節とは異形なので勝手に親戚の一人として認めてこなかったのですが、 DNAの塩基配列という最も基本的な形質で、スミレ属の中でChamaemelanium節の一員であることを主張しているのですから同意せざるをえませ ん。これはえらいことです。日本でもキバナノコマノツメ類としてV. bifloraとV. crassa(タカネスミレ)がいくつかの亜種や変種を伴って分布していますし、アジア全体では中国を中心にいっぺんに親戚がどっと増えてしまうことにな ります。またキバナノコマノツメより形態的にはもっとオオバキスミレに似ている謎の種V. alliariifolia(ジンヨウキスミレ)(2倍体)の所属も気になってきます。
さらにこの論文を読んで驚いたのは、スミレ属のそれぞれの種が系統樹上で分岐したおおよその年代を推定していることです。DNA分子の突然変異が
一定の
確率で起こることから、系統樹の何カ所かの分岐点の年代が化石や地史的なイベントなどで較正出来たならば、系統樹に時間の目盛りを入れる事が出来るという
わけです。
マルクーセンさん達は、その方法を使ってV. bifloraとV. brevistipulataが分岐したのはおよそ1300万年前、V.
unifloraが出現したのはおよそ1600万年前、V. biflora・V. brevistipulataの祖先種とV.
pubescens・V. glabellaの祖先種が分岐したのもおよそ1600万年前、V.
unifloraの祖先種と上記4種の祖先種が分岐したのはおよそ1800万年前と結論づけました。まだ相当の誤差を伴ってはいますが現生の種のDNAを
調べることでこのように過去のおよその分岐年代までが分かってしまうと言う、近年の分子系統学の進歩に全く驚いてしまいました。
私はこの論文を翻訳した後、その内容をより理解するために、分子系統学のイロハから勉強し始めました。葉緑体DNAを使った日本語の論文もいろいろ探し
出して読んでみました。
2000年に北海道大学図書刊行会から出た「高山植物の自然史」という本があります。第1章が植田邦彦・藤井紀行さんの「高山植物のたどった道」
という
論文でした。アリューシャン列島から北海道及び本州中部地方までのヨツバシオガマ24集団の葉緑体DNAを解析すると、東北以北と以南の二つの系統にはっ
きり分かれ、どうやら古い氷河期に本州まで南下したヨツバシオガマが間氷期に本州中部の高山帯をレフュージアとして残り、続く氷河期に東北地方にまで分布
を拡大。一方北方の新しいヨツバシオガマも北海道を通過して東北地方まで南下し、月山・飯豊山では両方の系統が生育していること、再度植物体を見直した
ら、両者の形態にいままで誰も気付かなかった違いがあったこと、両者が接して生育している所でも遺伝的に混じり合っていないことが分かったという驚くべき
内容でした。21世紀の生物学はDNAの時代だと心騒いだのをはっきりと覚えています。この研究は植田邦彦先生の指導の下、藤井紀行さんの博士研究として
なされたもので、1997年に博士論文となって金沢大学に提出されました。日本での葉緑体DNAによる分子系統学の草分け的な研究でした。
この「日本産高山植物の分子系統地理学的研究」と題された90ページに及ぶ博士論文は国立国会図書館のデジタル化資料としてインターネットに公開されて
いました。ありがたいことです。
マルクーセンさん達の論文と藤井紀行さんたちの論文には、共通点がありました。それは、解析に使用した植物のDNAのデータを国際的なデータベースに登
録してあるということです。アメリカ・ヨーロッパ・日本が共同で国際DNAデータバンクを作り、研究者は論文に発表した研究の証拠としてDNAをそこに登
録し、またそこから研究に必要なデータをダウンロードして比較検討することが出来ます。
とてもありがたいことに私のような部外者にもこのデータベースに
アクセスする道が開かれているのです。
マルクーセンさん達のViola属の登録データの中から、調べたいChamaemelanium節5種の葉緑体DNAデータと、さらにその中のV.
brevistipulataのデータを使って逆にデータベース全体から同じ領域の相同な近縁種のデータを検索して、韓国の植物学者の研究に使われたV.
brevistipulata var minor(ダイセンキスミレ)とV.
orientalis(キスミレ)の葉緑体DNAを見つけダウンロードしました。
これらV. brevistipulata(オオバキスミレ)に近縁な7種類と、外群としてV. serkirkii(ミヤマスミレ)とV.
langsdorfii(オオバタチツボスミレ)を合わせた9種類で自分でも見よう見まねで系統樹というものを作ってみました。
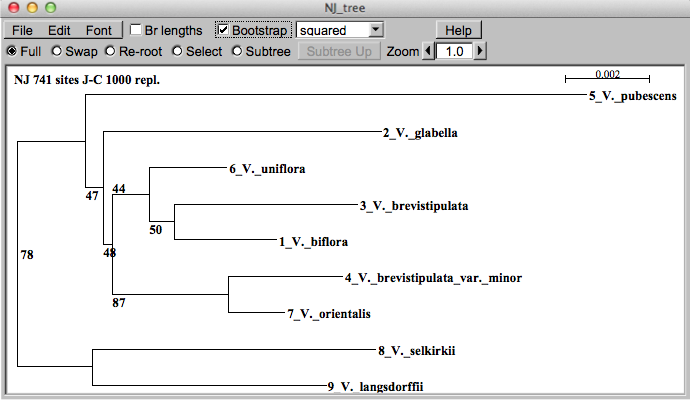
葉緑体DNAのtrnL-F領域にもとづいてSeaviewというフ
リーソフトを使って近隣結合法で作成した系統樹です。横枝の長さは塩基配列の異なる
度合を表しています。また枝の上の数字はその分岐が支持される確率を表しています。これを見ますとV. orientalisとV.
brevistipulata var minorの単系統が高い確率で支持されていて、V. brevistipulataはV.
biflora、V. unifloraと単系統となっています。この推定が事実ならばとても興味深いことです。このV. orientalisとV.
brevistipulata var
minorの単系統性は他のソフトを使っても、系統樹を作る方法を変えてもほぼ同じ結果が現れます。つまりダイセンキスミレは形態的にオオバキスミレの変
種ということになっていますが、DNAで見るとキスミレの方にはるかに近いということになります。このV. orientalisとV.
brevistipulata var
minorは韓国で採集されたものですが、やはりアジアのChamaemelanium節の全体像の解明にはもっと多くの材料を使った研究が現れるのを期
待するしかなさそうです。
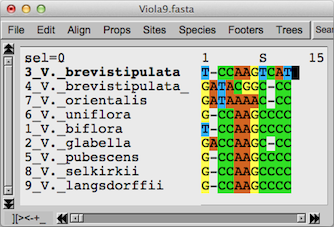
もう少し詳しくこの3種類について塩基配列を見てみます。上の図は9種類の塩
基配列を多重整列させてから両端を揃えた929サイトについて、3・4・7の3種類について塩基配列に違いのある部分11サイトのみを取り出した状態で
す。系統樹を作るときには挿入・欠失( –
のある行)を無視して作成しているのですが、ここではいっしょに考えてみます。例えば2行目のA(アデニン)が3ヶ所で見えているところは、上方の3種類
だけ見ればV. brevistipulataで欠失しているように見えますが、他の種の状態から考えてV. orientalisとV.
brevistipulata var minorの祖先種、及びV. glabellaで挿入があったと想像されます。V.
orientalisとV. brevistipulata var
minorでは929サイト中わずか3ヶ所の塩基が異なるだけです。それに対してV. brevistipulataとV.
brevistipulata var minorは10ヶ所の違いがあります。V. orientalisとV.
brevistipulataは9ヶ所の違いです。この違いが上記の系統樹に表現されています。
ここで藤井紀行さん達のヨツバシオガマの葉緑体DNAについても見てみます。研究では太平洋沿岸地帯全域から24集団から28個体を解析していま
すが、私はその中から、ウナラスカ島、大雪山、礼文島〜飯豊山、利尻島のデータ。これが東北以北のグループ。飯豊山〜雨飾山、白山、北岳のデータ。これが
東北以南のグループ。そして外群にエゾシオガマのデータをダウンロードしました。藤井さん達は葉緑体DNAの3ヶ所の領域合計1918サイトの解析を行っ
ているのですが、私が取得したのは上記のオオバキスミレ類の解析に使われた領域と重なる402サイトの部分のみです。8個体のみ、4分の1以下のサイトに
よる系統樹ですのでその分情報は少なくなるのですが、これでもやはり北方と南方の二つのクレードが分かれるのが見て取れました。
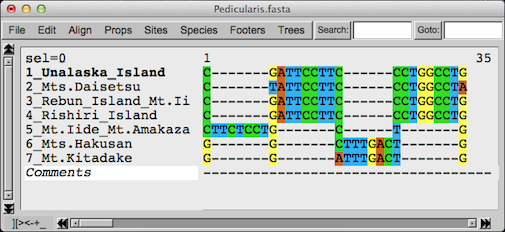
ここでオオバキスミレ類3種類のときと同じように、東北以北 (1〜4)と以南(5〜7)の二つのグループで塩基に違いのある部分のみを取り出して図にしてみました。そうすると内部での違いを置いておいて、グループ としての塩基配列の違いは挿入・欠失は2ヶ所、塩基置換がわずか1ヶ所だけです(7又は6塩基の挿入・欠失は元々連続していたものです)。藤井さん達もヨ ツバシオガマのDNAデータを使って北方系統と南方系統の 分岐年代を推定しています。結果は約769〜263万年前となりました。藤井さん達はこの値では第四紀より以前となってしまうので、かなり過大評価になっ ているのではないかと疑問符をつけておられました。
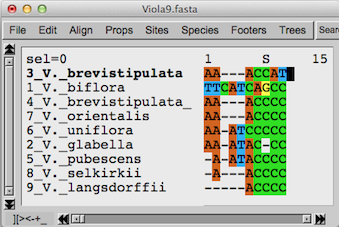
上の図はさきほどと同じオオバキスミレ類をSeaviewで多重整列させた
データから、3のV. brevistipulataと1のV. bifloraが異なる部分のみを取り出したものです。
マルクーセンさん達はV. brevistipulataとV.
bifloraが分岐したのはおよそ1300万年前としています。929サイトについてV. brevistipulataとV.
bifloraの塩基配列の違いは上の図のように10ヶ所です。従って違いが3ヶ所のV. orientalisとV. brevistipulata
var
minorが分岐したのはおよそ390万年前ということになります。その数字は藤井さん達のヨツバシオガマの両系統の塩基の違い、402サイトで3ヶ所で
約769〜263万年前という研究とそれほど違いはありません。
ヨツバシオガマとオオバキスミレ類の葉緑体DNAの同じ領域で進化速度に違いがないとすれば、キスミレとダイセンキスミレが遺伝的に分化したのは、ヨツ
バシオガマが二つの系統に分化したのと同じくらい前であったらしい。藤井さん達の書かれたようにこれは過大評価なのかもしれませんが、どうやら最後の氷河
期の数万年というような短い期間の出来事ではないことは確かなようです。
Viola
guadalupensisというスミレはアメリカテキサス州のグアダルーペ山脈で800万年ほど昔に生まれ、繁栄し、そして現在は石灰岩の岩場数カ所に
約750株を数えるのみだと言います。Viola brevistipulata var minorやViola
brevistipulataも同じように、はるか昔に生まれ、繁栄し、今はあちこちに分断されて生育しています。悠久の命の営みの不思議に想いを馳せま
す。
|
|
|
|